Los fánnidos son una familia pequeña de dípteros con calípteros cuyas especies miden, en general, de 3 a 5 mm de longitud. Esta familia está distribuida por todo el mundo y se han descrito más de 360 especies, 85 de ellas se encuentran en Europa. Muchas de ellas son abundantes y presentan una distribución amplia en las zonas más templadas de la región paleártica.
Se clasifican en 5 géneros:
Se clasifican en 5 géneros:
- Género Euryomma Stein, 1899
- Género Piezura Rondani, 1866
- Género Fannia Robineau-Desvoidy, 1830
- Género Australofannia
- Género Zaelandofannia
Los Fanniidae son básicamente habitantes de los bosques y son relativamente raros en paisajes abiertos y humedales.
Los machos de casi todas las especies forman enjambres bajo las ramas de los árboles y en los caminos forestales, con individuos siguiendo trayectorias horizontales definidas y con frecuencia entrando en combate con otros individuos. El significado exacto de esta actividad no está aún claro. Las hembras se encuentran en la vegetación del suelo siendo atraídas por la materia orgánica en descomposición y los excrementos, aunque hay algunas especies, denominadas secretófagas, que atacan al ganado en los pastos, así como las personas que sudan en verano.
La mayoría de las larvas se desarrollan en materia orgánica descompuesta de muchos tipos, aunque sobre todo vegetal; son fácilmente reconocibles por presentar el cuerpo aplanado con procesos laterales ramificados.
Algunas de las especies de la familia Fanniidae son indicadores útiles en entomología forense.
Hasta principios de los años 70 esta familia se consideró una subfamilia o tribu de los Muscidae. No obstante, los Fanniidae se separan fácilmente de ésta por :
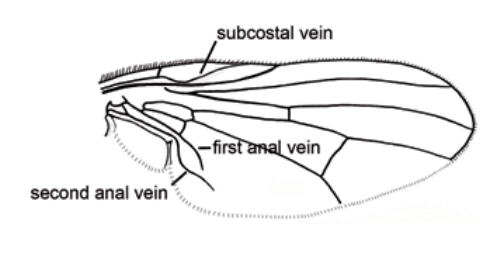
1) presentar la vena subcostal acodada cerca de la base y una primera vena anal muy corta pero cuya prolongación imaginaria cruzaría a la segunda vena anal.
2) tener en la tibia posterior unicamente una cerda dorsal por debajo de la mitad de la tibia + una cerda dorsal preapical.

3) En los machos, la tibia media tiene una pubescencia erecta por debajo; en las hembras, la frontalia no tiene cerdas cruzadas, las órbitas frontales son anchas y convexas hacia la línea media de frons, y hay dos pares de cerdas orbitales superiores fuertes, con el par anterior girado hacia afuera.
Morfologia de los individuos adultos de la familia Fanidae:
Los machos tienen la cabeza semicircular y cóncava por detrás, las hembras la tienen ovoide y convexa por detrás. Los machos son holópticos y las hembras del género Fannia, ambos sexos de las especies de Euryomma, Australofannia Pont, Zealandofannia, algunos Piezura Rondani tienen la cabeza dicóptica. La mayoría de los ojos están desnudos, pero hay especies que los tienen escasamente setulosos. Los ojos están delimitados por una placa fronto-orbital relativamente estrecha o ancha. La placa fronto-orbital y el frontal vitta (interfrontales) ocupan la línea central de la cabeza, entre el triángulo ocellar y la lunula. En los machos del género Fannia, esta área es generalmente estrecha o muy estrecha, de modo que los ojos parecen casi tocarse en parte de su longitud. Por lo general, las placas fronto-orbitales femeninas tienen un margen mediano interno convexo, que es un carácter automomórfico de la familia. Las placas fronto-orbitales en machos y hembras tienen una fila de setas frontales que varían en número, longitud y fuerza. Las hembras del género Fannia y ambos sexos de Australofannia, Zealandofannnia, Euryomma y algunos Piezura tienen 1 reclinación superior y 1 seta orbital de exclusión inferior (autofomorfia), así como un número variable de filas de setas a lo largo de toda la longitud de las placas fronto-orbitales. Los machos del grupo canicularis del género Fannia y algunas especies de Piezura tienen la seta orbital superior (reclinada) fuertemente desarrollada. El triángulo ocelar en el vértice es muy distinto y los tres ocelos están desarrollados. A cada lado del triángulo ocular se encuentran las setas verticales externa e interna, mejor desarrolladas en hembras, seguidas de una fila de setulas postoculares que generalmente son simples, pero pueden tener una disposición doble o irregular. La cara es la parte de la cabeza, debajo de la antena. Está separado de los márgenes oculares por un parafacial. El parafacial está generalmente desnudo, pero puede tener filas de setulas (como en las hembras de los grupos de canicularis) o raramente una fila de setas. En el lado de la parte inferior de la cara, un par de setas fuertes y generalmente cruzadas están presentes en el ángulo de de las vibrisas. La gena es generalmente angosta, con el ancho por debajo del margen inferior ocular del menos del doble del tamaño del postpedicelo, pero puede ser amplio.

La antena consta de dos segmentos basales cortos (escapo y pedicelo) y un tercero más largo, el postpedicelo, que casi siempre es más corto que la cara. La antena puede variar en color de negro o marrón a amarillo. El postpedicelo lleva una arista en posición dorsal habitualmente. La arista suele estar desnuda o bien tener una corta pubescencia, pero en ocasiones es plumosa, como en el género Piezura, o subplumosa, como en Australofannia y en el grupo de Fannia admirabilis de Sudamérica. Las partes bucales consisten en un tipo de probóscide de succión. La parte tubular corta de la probóscide se forma a partir del labio esclerotizado. El prementum puede brillar o ser pruinoso en el lado ventral. La parte apical de la probóscide termina en dos labelas. Los palpos maxilares suelen estar bien desarrollados, pero pueden tener un tamaño muy reducido y pueden ser filiformes o de forma espatulada. Varían en color de marrón oscuro a amarillo.
El tórax es como en la mayoría de los dípteros que tienen caliptero. El pospronotum está bien desarrollado y está estrechamente asociado con el mesonotum (scutum); Sus márgenes posterolaterales forman los lóbulos postpronotales. Este lóbulo en Fanniidae tiene tres setas (dos fuertes y una más débil) y un número variable de setulas. Lateralmente, el propleurón se divide en una porción anterior, el proepisternum, y una porción posterior más pequeña, el proepimeron. Estos dos escleritos forman un arco sobre la coxa anterior, que se sitúa debajo del espiráculo anterior. El proepisternum tiene 2 setas y el proepimeron 1 seta, cada uno con un número variable de setulas. El scutum está provisto de un número variable de filas longitudinales de setulas acrosticales. El número de filas suele ser constante en los diferentes grupos de especies. Cinco pares de setas dorsocentrales, dos delante y tres detrás de la sutura transversal, son un carácter de familia. Solo Fannia abnormis carece de las setas dorsocentrales presuturales y en Euryomma y Zealandofannia se reduce la dorsocentral presutural anterior. Lateral a las filas dorsocentrales se encuentran las setas intraalares, de las cuales usualmente hay un par presutural y dos postsuturales, pero solo hay un par postutural en Zealandofannia. Las setas postalar y supraalar (presutural y postutural) suelen ser distintas. Detrás del margen lateral de la sutura transversal y en la región anteromedial del proceso alar notal se encuentra el área prealar. Aquí están las setulas prealares, que son de gran importancia toxonómica. El nombre "prealar" fue introducido por Stein para designar el primer y segundo setae/setas supraalares inmediatamente detrás de la sutura. En Fanniidae las setulas pueden estar ausentes, reducidas o duplicadas. Su posición también puede variar, ya que pueden estar más cerca de la seta supraalar o de la sutura. El escutelo, situado posteriormente al escutum, tiene dos pares de setas fuertes y uno o dos pares de setas finas marginales, un par de setas discales antes de la punta y unas pocas a varias setulas discales adicionales. Los márgenes laterales debajo de las setas fuertes y la superficie ventral son lampiños. El mesopleurón se divide por la sutura pleural en un episterno anterior y un epimeron posterior. El episterno se divide transversalmente por una sutura en un área superior, el anepisternum, y un área inferior, el katepisternum. Las setulas anepisternales cubren la mayor parte del anepisternum, y generalmente forman una fila de setas a lo largo de su margen posterior. El katepisternum generalmente tiene dos setas fuertes además de algunas setulas finas, pero hay una segunda seta posterior (más baja) en Zealandofannia. Del mismo modo, el epimeron se divide transversalmente en un anepimeron y un katepimeron, que siempre están desnudos en Fanniidae. Debajo de ellos está el meron, que también está desnudo en Fanniidae.


El ala tiene una venación visiblemente uniforme en prácticamente todas las especies. La forma de la subcosta y de la primera y segunda venas anales son apomorfías de la familia. La subcosta corre recta en su mitad apical, sin ninguna curva sinuosa. La primera vena anal se acorta notablemente, y la segunda vena anal es fina y tiene forma de arco. En Euryomma, la segunda vena anal es más larga, pero su extensión imaginaria todavía interceptaría una extensión imaginaria de la primera vena anal antes del margen del ala. Las alas pueden variar en color, desde translúcidas hasta muy ahumadas. La membrana está siempre completamente cubierta con microtrichia minúscula. Los calipteros son en su mayoría del mismo tamaño, por lo que el margen externo de la parte inferior es claramente visible. Sin embargo, la parte inferior del caliptero puede variar en su forma, desde redondeado como en el grupo de Euryomma y Fannia canicularis, hasta ligeramente ovalado como en la mayoría de las especies de Fannia; o puede ser muy estrecho y casi como una franja, como en Australofannia y el grupo de Fannia serena, o alargado y en forma de lengua, como en Fannia australis Malloch. Los calipteros son generalmente blancos, pero pueden ser marrones en el margen exterior. Los halterios son en su mayoría de color blanco amarillento, pero son de color marrón a negro en los machos de algunos grupos.
Las patas proporcionan una serie de valiosos caracteres diagnósticos, con algunas modificaciones estructurales sorprendentes en los machos, así como disposiciones variables de setas y pelos.
El fémur anterior puede ser delgado o estar como hinchado, generalmente desnudo en la superficie anterior y con un número variable de filas de setas en las superficies posterodorsal, posterior y posteroventral. La tibia delantera tiene una seta posterodorsal preapical y setae apical ventral y en la superficie posteroventral. Algunas especies tienen una seta corta o una fila de setas cortas en la superficie anterodorsal. Algunos machos tienen una seta posterior submediana, o setas finas en la superficie posteroventral. Los tarsómeros delanteros suelen ser oscuros y de forma normal, pero pueden ser cambiar en forma y color en unos pocos machos, como en el grupo Fannia anthracina, donde son aplanados y ensanchados y parcialmente a totalmente blanquecinos. En los machos de unas pocas especies están adornados con pelos finos.
El fémur medio en los machos no muestra una gran variación en la forma, aunque puede estar ligeramente curvado o indentado en la parte apical. Suelen estar presentes hileras distintas de setas anteroventral y posteroventral que pueden estar engrosadas y / o duplicadas en los machos. A veces se forma un peine de setulas preapical. En los machos, el ensanchamiento de la tibia media en su porción apical (excepto en el grupo de Fannia canicularis) y la pubescencia en la superficie ventral son apomorfías para la familia. La tibia media masculina también puede tener un par de indentaciones en su mitad basal, como en el grupo de Fannia anthracina. La disposición de las setas es bastante uniforme, pero las setas anteriores, anterodorsales y posterodorsales pueden duplicarse o triplicarse en algunas especies. Las hembras de algunas especies tienen setas anteroventral y posteroventral. En los tarsómeros medios, el tarsómero basal puede estar armado en algunos machos con una espina ventral en forma de gancho o en forma de varilla o un grupo de setulas cortas y densas que forman una cresta en su base.
La coxa trasera está desnuda o tiene 1–3 pelos cortos en su margen interno posterior. El fémur posterior puede variar de delgado a ensanchado y puede tener ensanchamiento ventral preapical. Las filas de setas anteroventral y posteroventral en los machos pueden desarrollarse o reducirse en una medida variable, o pueden estar completamente ausentes. La tibia trasera tiene una seta submediana en la superficie dorsal, en línea con la seta dorsal apical, que es una apomorfia para la familia. Existen diferentes números de setas y series anterodorsales, anteroventral y en ocasiones también posteroventrales. Algunas setas son ocasionalmente alargadas y parecidas a pelos o se agrupan de varias maneras en los machos.
El abdomen es generalmente de estructura uniforme, excepto en los segmentos terminales. Los cinco segmentos principales son visibles en una vista dorsal . En los machos el abdomen es más amplio a nivel del segmento 2, excepto en el subgrupo de mollissima de Fannia en el que el abdomen es mas ancho a nivel de segmento 4. Un patrón más o menos distinto suele estar presente en la superficie dorsal pruinosa del abdomen masculino, en forma de una línea media oscura o marcas triangulares oscuras en cada tergito, rara vez de parches laterales simétricos. El abdomen de la hembra suele ser uniformemente oscuro, con algunas excepciones. El esternito 1 es ancho y densamente setuloso, especialmente en los machos. El esternito 5 difiere de los esternitos anteriores, y generalmente se amplía y tiene setas más fuertes a lo largo del margen posterior, además de que a menudo tiene otras modificaciones características de especie.
La terminalia masculina incluye los segmentos postabdominales y el hipopigio. Los cerci se fusionan en una placa mediana que es extremadamente variable y con frecuencia tiene una forma específica para cada especie. El epandrio es de forma hemisférica y tiene un par de surstilos en su margen posteroapical. Los surstyli son frecuentemente de forma característica para la especie y pueden ser robustos, delgados, doblados, bífidos, etc., pero son bastante uniformes en el aspecto en el grupo de Fannia canicularis. El hipandrio es semicircular, conectado con la base edeagal por medio de un puente esclerotizado. El pregonio (gonópodo) está ausente en la familia, y el estrecho postgonio (paramere) está intimamente unido al falo. El falo en sí es en su mayoría membranoso, más esclerotizado en algunas especies (Piezura, Euryomma y algunas especies de Fannia).
La terminalia femenina tiene la forma de un oviscapto alargado y telescópico, sin estructuras especializadas; solo en Australofannia y Fannia capitalis Pont (Australia) se acorta notablemente el oviscapto. Consiste en los segmentos abdominales del 6º al 9º, enfundados dentro del segmento 5º. Las tergitas y esternitas están esclerotizadas en una extensión variable. El cerco y las dos placas anales (el epiprocto dorsal y el hipoprocto ventral) son en su mayoría simples, cubiertos con finas setulas. El hipoprocto tiene un par de lingulas cortas en cada vértice anterior. Algunos caracteres distintivos derivan de la posición de los dos pares de espiráculos postabdominales,y del número y forma de las espermatecas, que pueden ser dos o tres, esféricas, en forma de pera o de forma alargada, con una superficie lisa o esculpida.
De la morfología de la larva pondré solamente un esquema:

El género Fannnia Robineau-Desvoidy, 1830
El género fue originalmente descrito por el entomólogo francés Jean-Baptiste Robineau-Desvoidy en 1830. Previamente varias de las especies de este género se colocaban dentro del género Musca. La especie tipo es Fannia saltatrix Robineau-Desvoidy, 1830 (= Musca scalaris Fabricius, 1794).
Es el género de mayor tamaño de la familia Fanniidae y se encuentra en todas las regiones zoogeográficas del mundo. Comprende más de 285 especies, la mayoría de las cuales son de la Región Holarctica.
La importancia médica e higiénica de las especies englobadas en este género, tales como Fannia canicularis (Linnaeus), Fannia femoralis (Stein), Fannia incisurata (Zetterstedt), Fannia pusio (Wiedemann) y Fannia scalaris (Fabricius), es bien conocida. Fannia canicularis y F. scalaris se han criado sin problema a partir de una variedad de materiales orgánicos en descomposición. Además, las larvas de F. scalaris se encuentran con frecuencia en pozos, letrinas y estiércol, y también se han criado junto con F. canicularis a partir de heces humanas. Algunas de las especies más abundantes se ven regularmente en corrales agrícolas utilizados para la cría de cerdos, vacas, caballos y aves de corral, y en granjas de pieles, donde aparentemente las larvas se desarrollan en excrementos de animales.
Diagnóstico del género:
Primera seta dorsocentral presutural mide más de una vez y media la longitud de la segunda. Seta katepisternal inferior ausente. Tibia media del macho generalmente ensanchada apicalmente. Ala con la segunda vena anal (A1 + CuA2) fuertemente curvada; su prolongación imaginaria intersectaría la primera vena anal (A2) mucho antes del margen del ala. Terminalia masculina generalmente con esclerito baciliforme distinto; Oviscapto de la hembra con 2 o 3 espermatecas.
En la Península Ibérica podemos encontrarnos las siguientes especies del género Fannia:
- Fannia aequilineata
- Fannia alpina
- Fannia armata
- Fannia canicularis
- Fannia carbonaria
- Fannia clara
- Fannia coracina
- Fannia corvina
- Fannia cothurnata
- Fannia difficilis
- Fannia fuscula
- Fannia glaucescens
- Fannia incisurata
- Fannia krimensis
- Fannia latipalpis
- Fannia lepida
- Fannia leucosticta
- Fannia manicata
- Fannia mollissima
- Fannia monilis
- Fannia norvegica
- Fannia ornata
- Fannia pauli
- Fannia polychaeta
- Fannia postica
- Fannia pseudonorvegica
- Fannia pusio
- Fannia rondanii
- Fannia scalaris
- Fannia serena
- Fannia sociella
- Fannia tunisiae
- Fannia vesparia
Se puede encontrar una Clave dicotómica para las especies europeas de esta familia en el siguiente artículo: Clave dicotómica para las especies europeas en Fanniidae (Diptera): new synonym, new records and an updated key to males of European species of Fannia ZooKeys 593: 91–115 (2016) doi: 10.3897/zookeys.593.7735
Fannia es, con mucho, el género más grande de la Familia Faniidae e históricamente hablando se ha dividido en una serie de grupos (Chillcott 1961; Rozkošný et al. 1997; Domínguez & Roig-Juñent 2008). Hennig (1955) y Chillcott (1960), basándose fundamentalmente en las terminalias de los machos propusieron varios grupos, algunos de ellos divididos en subgrupos, pero solamente uno de ellos se ha demostrado monofilético (grupo antracina). El grupo canicularis es otro de los grupos clásicos de Hennig y del que él enumeró los siguientes caracteres de grupo: seta orbital superior desarrollada más o menos claramente en el macho, coxa posterior con 1 o más setulas posteroventrales y surstylus masculino que se extiende desde el margen interno del tergito abdominal 9. Chillcott sugirió que este grupo era primitivo y estaba más estrechamente relacionado con los géneros Euryomma Stein, 1899 y Piezura Rondani, 1866 que con otros grupos de especies de Fannia. Usando los caracteres morfológicos y las estructuras genitales, dividió este grupo en 3 subgrupos, el subgrupo F. canicularis, el subgrupo F. pusio (= subgrupo F. leucosticta) y el subgrupo F. glaucescens. Esta clasificación ha sido aceptada y seguida por muchos especialistas.
Diagóstico del grupo F. canicularis:
El caliptero inferior se proyecta claramente más allá del superior. La coxa media del macho carece de espinas en forma de gancho. Coxa posterior con algunas pequeñas setas en el margen posterior interno (rara vez ausentes). Tibia posterior de la mayoría de las especies con una fila de pelos que incluye 1 o 2 pelos potentes. Katepisternum masculino sin espinas en el margen inferior; generalmente es pequeño, parecido a un vello, pero si es fuerte, entonces el proepisternum con pelos. Cerci masculino de forma triangular, rara vez rectangular, surstyli con 2 ramas, la anterior usualmente con pelos, pero si los surstyli no están ramificados, estas estructuras son cortas, amplias y muy escleroizadas. Algunos machos con setas robustas parafaciales, y hembras con setas o setulas parafaciales superiores.
A continuación pongo una clave dicotómica para la diferenciación entre los tres subgrupos de F. canicularis:
1. Abdominal tergites without lateral markings, only with median longitudinal stripes or triangular markings, or even lacking markings………………………………………………………………….2
- Abdominal tergites 3 and 4 with median longitudinal stripes and lateral spots ……………………………………………………………………………….F. leucosticta-subgroup
2. At least 1 surface (including av, ad, pd, or pv surface) of hind tibia with 2 or more setae, but if only with 1 seta then abdominal tergites usually with triangular markings, male cerci gradually constricted towards apex, triangle-like, surstyli with 2 branches, anterior one usually with hairs…………………………………………………………………………F. canicularis-subgroup
- Each surface (including av, ad, pd, or pv surface) of hind tibia usually with only 1 seta, but if with 2 setae then parafacial with strong setae; abdominal tergites with or without median longitudinal stripes; male cerci rectangle-like, surstyli heavily sclerotized, broad, and short, and unbranched…………………………………………………………………….F. lucidula-subgroup
Fannia canicularis
La palabra Fannia deriva del griego “phanos, -e, -on” que significa, claro o brillante. Canicularis deriva del latín “canicularis, -e” que significa, canicula, palabra que a su vez deriva de {can}, es decir, perro, y tiene relación con la constelación de Sirio, llamada “la perrita”. La salida de Sirio coincide con los primeros días de agosto.
Esta mosca es ligeramente más pequeña que la mosca doméstica común, y aparecen también en las viviendas, normalmente alrededor de los focos luminosos, con un vuelo horizontal en zigzag, que es típico de la especie.
Fannia canicularis es la vulgarmente denominada mosca doméstica menor, para ponerla en contraposición con la mosca doméstica común, que es de mayor tamaño. Fannia canicularis mide 4 - 6 mm de longitud. El adulto es de color gris-negruzco, con tres rayas negras difusas dispuestas longitudinalmente sobre el dorso del tórax; estas lineas son mucho mas definidas y nítidas en el macho que en la hembra. Los lados del tórax son de color algo más claro. Los halterios son amarillos. La cabeza es gris, con rayas frontales negras y lados grises. Los machos se diferencian de las hembras porque son holopticos, mientras estas son dicópticas. Macho con una clara seta orbital reclinada. Hembra con setulas en placa fronto-orbital continua abajo a parafacial. Las antenas, palpus y la mayoría de las patas negras. Las antenas son similares a las de la mosca doméstica común, pero la arista respectiva es delgada y aparece cubierta de una fina pubescencia. Los dos primeros segmentos del abdomen son de color amarillo translúcido con un color basal marrón oscuro. Las oscuras marcas trapezoidales de los machos son difícilmente reconocibles en las hembras. La coxa posterior posee 2 setulas en la superficie postero-apical. Solo hay 1 setula adyacente a la seta proepimeral. La tibia trasera tiene varias setas cortas por encima y por debajo de la seta anterodorsal fuerte.. Tibia media de estructura simple, con pubescencia ventral corta y fina.

Hembra de Fannia canicularis
Descripción detalallada de ambos sexos.
El macho tiene una longitud corporal de 4.5–7.0 mm. Su cabeza tiene una frons estrecha de color marrón oscuro a negro y en el punto más angosto es tan ancha como el ocelo anterior. La placa fronto-orbital es plateada y pruinosa,. El par más alto de las setas frontales mide 0,9 x ancho del ocelo anterior. Hay hasta 13 pares de estas largas setas frontales. Hay una seta orbital superior que es reclinada. El ojo es lampiño con las facetas internas superiores más grandes que las restantes. Hay una fila de setulas postoculares superiores. El parafacial es pruinoso. La cara de color gris claro. El parafacial en la base del postpedicelo mide 0.6 × ancho del postpedicelo y es lampiño. La gena es de color gris oscuro a negro. El escapo y el pedicelo son negros, al igual que el postpedicelo que mide de largo dos veces el ancho y está cubierto de pruinosidad gris. La arista es negra, casi lampiña. El prementum de la probóscide es de color parduzco pruinoso. Los palpos son negros, rectos, de ápice tan ancho como la base. El tórax tiene un color de fondo negro. El Scutum es de color gris a marrón grisáceo pruinoso, con color de fondo marrón oscuro extendiéndose a lo largo de las líneas acrostical, dorsocentral e intraalar desde el cuello hasta casi el escutelo. El lóbulo postpronotal tiene 3 setas, la interna débil y 1-2 prealares muy cortas cerca de la supraalar. Hay 2 setas proepimerales sin ninguna otra seta adyacente. Las patas son de color marrón oscuro a negro, con las rodillas amarillas. La tibia delantera es amarilla en el cuarto basal. Proporción largo/ancho del fémur anterior = 6 y con 1 hilera de setas posterodorsales, 2 filas de setas posteriores, 1 posteroventral basal y una fila posteroventral en los ¾ de la zona apical. La tibia anterior tiene 1 seta dorsal preapical fuerte, 1 apical ventral, posteroventral y anterodorsal apical muy corta y 2 anterodorsales cortas en la mitad apical. El tarsomero anterior mide 1 0.5 × longitud de la tibia. Los tarsómeros 4 y 5 son cortos y tan anchos como largos. El fémur medio tiene una fila completa de setas anterodorsales, 2 filas de setas anteriores muy cortas, una fila anteroventral corta y fina en el tercio basal, que va haciéndose más robusta y con forma de peine en el tercio apical, una fila posteroventral fuerte, tan larga como la anchura femoral, que se vuelve corta, robusta y con forma de peine en el cuarto apical y fila posterior fina en la base, cada vez más larga y fuerte y con 5-6 pelos más fuertes en el cuarto apical. La tibia media es simple en su estructura con pubescencia ventral muy corta, 0.25 × ancho de la tibia, con 1 seta anterodorsal y 1 otra posterodorsal en la mitad apical y setas anterior y anteroventrales fuertes, o en su defecto con setas débiles y cortas en el ápice. La coxa posterior tiene 2 setulas en el ápice de la superficie postero-apical. El fémur posterior es simple en su estructura, con un indice largo/ancho = 6.25, con una fila de setas anterodorsales, de las cuales las 4-5 son apicales con dirección dorsal, una fila de setas anteroventrales muy cortas, solo 3 a 4 pelos en el tercio apical tan largos como el ancho femoral y una superficie posteroventral con solo setas finas cortas, del tamaño de la mitad del ancho femoral. La tibia trasera tiene setas submediales y dorsales preapicales, 1 mediana anterodorsal fuerte y 3–8 setas cortas superiores e inferiores a esta. Hay una seta anterodorsal apical bien desarrollada, 2–3 setas anteroventrales medias mas una seta anteroventral apical. El ápice de las superficies ventral y posteroventral posee un corto ctenidio y una corta seta apical posteroventral. Las alas son hialinas con venas de color marrón amarillento. La basicosta es de color amarillo, y la tégula de color naranja a marrón. Los calypteros son de color blanco, blanco crema en los márgenes, siendo el inferior mas largo que el superior. La cabeza del caliptero es de color blanco amarillento y la base de color marrón claro. El abdomen es alargado y con color de fondo negro, syntergite 1 + 2 y tergitos 3–4 de ancho uniforme.pero de color amarillento, especialmente en los lados; raramente son completamente negros. Los tergitos son de color gris claro pruinoso, con un color de fondo negro y a lo largo de todos los tergitos; las áreas oscuras se expanden hacia atrás en los sintergita 1 + 2 y los tergitos 3–4 para formar marcas triangulares, a veces menos definidas en el tergito 4. El esternito 1 es densamente setuloso. El margen posterior del esternito 5 es recto, con las setas formando una línea longitudinal central. El hipandrio es ancho, está fuertemente esclerotizado ysus brazos hipandriales están dirigidos hacia adentro. Los parmeres tienen forma de cuerno, están fuertemente esclerotizados y se hayan rodeando el aedeagus membranoso. La placa cercal es pequeña y está fusionada. El esclerito baciliforme está ausente. El surstylus está formado por un proceso principal aplanado, de lados paralelos y un proceso ventral, lateral, más pequeño y deprimido que lleva un grupo de setulas en el ápice. Cada surstylus está directamente y ampliamente conectado al hipandrio.



La hembra tiene una longitud corporal de 4.8–5.2 mm. En la cabeza la frons y frontal vitta son anchas, siendo la distancia entre los ojos en la parte media de la cabeza de 1/3 del ancho de la misma en este punto. La placa fronto-orbital es de color gris pruinoso. El triángulo frontal es muy corto, no alcanza el nivel de la seta orbital inferior. Hay 7-8 pares de setas frontales, de las cuales 3 son fuertes. La seta orbital inferior se haya más o menos a medio camino entre el margen ocular y el margen de la placa fronto-orbital o ligeramente más cerca de esta última. Las setulae fronto-orbtales estan en varias filas por encima, en una fila por debajo y continúan distalmente más allá del nivel de la base de postpedicelo. El tórax es de color marrón generalmente más amplio y más difuso. Las patas tienen el mismo color que las de el macho, con trocánter posterior amarillo. El fémur medio posee algunas setas anteroventrales cortas y finas en el cuarto basal, 1 seta posteroventral en la base, o de lo contrario solamente setulas en las superficies ventrales. La tibia media no posee pubescencia ventral. El fémur trasero está ventralmente lampiño, excepto 2–3 setas fuertes anteroventrales en la zona apical. El abdomen tiene forma de corazón, siendo el tergito 4 aproximadamente la mitad de ancho que el ápice del tergito 3. El color de fondo del abdomen es fundamentalmente oscuro, con algo de amarillo en la base y, a menudo, también en los lados de los tergitos 3 y 4. Todos los tergitos son pruinosos de color gris con trazas medias oscuras visible sen todos los tergitos. Los cerci son normales, cortos. El hipoprocto es ligeramente más ancho que largo y está cubierto uniformemente de setas. El esternito 8 se haya reducido a un par anterior de placas cubiertas con setulas y dos placas posteriores lampiñas ligeramente más grandes que las placas anteriores. Los espiráculos postabdominales 7 y 8 se hayan situados en el tergito 6. Las dos espermatecas son esféricas y redondeadas, de superficie blanda y con conductos no esclerotizados.


Las hembras realizan las puestas en lotes de hasta 50 huevos, en lugares húmedos y en material en descomposición, pudiendo tener varias generaciones por año. En total pueden poner hasta los 2000 huevos. Los huevos de la Fannia canicularis son blancos, con un par de rebordes o aletas longitudinales dorsales. Pueden flotar en materia orgánica líquida y semilíquida en descomposición, especialmente de aves de corral, heces de vacas y perros, desechos de cocina como patatas o zanahorias podridas, ensilado, compost queso, tocino y pescado seco. Se encuentran comúnmente en depósitos de basura, contenedores con ruedas, camiones de basura y otros lugares donde se almacenan los desechos de alimentos. Eclosionan en 20-48 horas a 24-27ºC y las larvas requieren 6 o más días para llegar a la pupación, que dura 7 o más días. El ciclo biológico completo (de huevo a adulto) es de 15-30 días, según la temperatura. A cualquier temperatura dada, el ciclo biológico es ligeramente más largo que el de la mosca doméstica. Tienen una esperanza de vida de dos a tres semanas. En Europa Central, alrededor de siete generaciones pueden desarrollarse por año. Las larvas son grandes consumidoras de material putrefacto, incluida carroña, contribuyendo a su reciclado.
Entre los Fanniidae, esta especie es la más frecuentemente asociada con la miasis urogenital e intestinal y debido a su oscilación entre excremento y alimento humano, se consideran posibles portadores de enfermedades.
En la naturaleza enjambran bajo las ramas de los árboles donde se las puede ver con su característico patrón de vuelo cambiando bruscamente de dirección, De mayo a octubre, entra con frecuencia en los edificios donde realiza su vuelo de patrulla, en el cual los machos supervisan, si es necesario, su distrito y atacan a los intrusos. Durante los descansos breves y en las horas nocturnas, las moscas se sientan en lámparas o en las paredes , dejando sus pequeñas marcas de excremento.




























